Moluscos Fósseis do Acervo

Trigonia sp 
Trigonia sp 
Trigonia sp 
Trigonia sp 
Trigonia sp 
bivalve 
bivalve 
bivalve 
bivalve 
bivalve 
bivalve 
bivalve 
bivalve 
gastrópode 
gastrópode 
gastrópode 
gastrópode 
Amonites 
Cearatites 
cefalópode amonóide 
cefalópode amonóide 
Orthoceras sp. 
Orthoceras sp. 
molde de conchas 
molde de conchas 
Coquina 
Ceratites 
cefalópode amonóide 
gastrópode 
gastrópode 
gastrópode molde interno 
gastrópode molde interno 
gastropode (molde interno)
Introdução
Depois dos artrópodes, o Filo Mollusca é o segundo maior grupo de animais em diversidade, sendo conhecidas 100.000 espécies viventes e 30.000 espécies fósseis. A grande variedade de formas e hábitos de vida desenvolvidos pelas diferentes classes deixou um riquíssimo registro fossilífero Fanerozoico, especialmente em sedimentos marinhos, e de maneira menos abundante em rochas sedimentares de ambientes continentais aquáticos ou mesmo terrestres.
Resultados de estudos moleculares indicaram a divergência dos mais antigos representantes do filo em rochas com até 1.2 bilhões de anos de idade. No entanto, as mais antigas raízes filéticas representadas por restos ou vestígios fósseis, são encontradas em rochas do final do éon Proterozoico, com idades entre 600 e 541 milhões de aos atrás, quando a mineralização de tecidos começou a dar os primeiros passos entre os metazoários.
Kimberella quadrata, encontrada em rochas da Austrália e Rússia com idades entre 558 e 555 milhões de anos, ainda no último período do Éon Proterozoico é, atualmente, o candidato ao mais antigo representante de Mollusca. Kimberella viveu em mares de águas rasas e oxigenadas, alimentando-se de cianobactérias e outros microorganismos componentes das camadas microbianas não deixou partes rígidas como conchas e outras estruturas preservadas. Porém, icnofósseis (Radulichnus) encontrados associados às marcas de descanso do seu corpo mole, são indicativas da presença de rádula, caráter autapomórfico dos moluscos.
Figura 6.1. Kinberella quadrata, Período Ediacarano, representação em vida, e Radulichnus, icnofósseis atribuídos à atividade da rádula.
Diferente da maiora dos filos, os moluscos deixaram as águas marinhas para colonizar água doce e ambientes terrestres úmidos.
A grande variedade de formas deve-se, principalmente, aos diferentes hábitos alimentares desenvolvidos. Os bivalves, representados principalmente por formas detritívoras e filtradoras da infauna rasa durante a Era Paleozoica, tornaram-se durante o Mesozoico a fauna conchífera dominante nos ambientes marinhos, especialmente pelo fato de terem ocupado de modo eficiente o interior do substrato. Além disso, monoplacóforos e gastrópodes possuem a rádula, aparelho raspador que possibilitou o surgimento de formas herbívoras, carnívoras e mesmo necrófagas. Os cefalópodes, também dotados de rádula, adquiriram mandíbulas fortes, tornando-se predadores e excelentes nadadores. Durante o Mesozoico, os cefalópodes experimentaram grande diversificação, mas foram quase extintos durante a crise do limite Cretáceo-Paleógeno, sendo representados hoje por poucos gêneros viventes.
Os moluscos fósseis representam organismos especialmente importantes do ponto de vista paleoecológico, uma vez que a maioria dos grupos tem representantes vivos, permitindo a utilização do princípio do atualismo na interpretação de sua morfologia funcional. As diferentes formas e hábitos de vida dos moluscos proporcionam dados importantes na reconstituição de ambientes antigos.
Embora a maioria das características morfológicas que definam as relações de parentesco entre as diferentes classes de moluscos sejam parte moles de difícil preservação no registro, a sobrevivência de diversos grupos ancestrais, a filogenia é bem resolvida quando considerada a maioria dos grupos fósseis conhecidos.
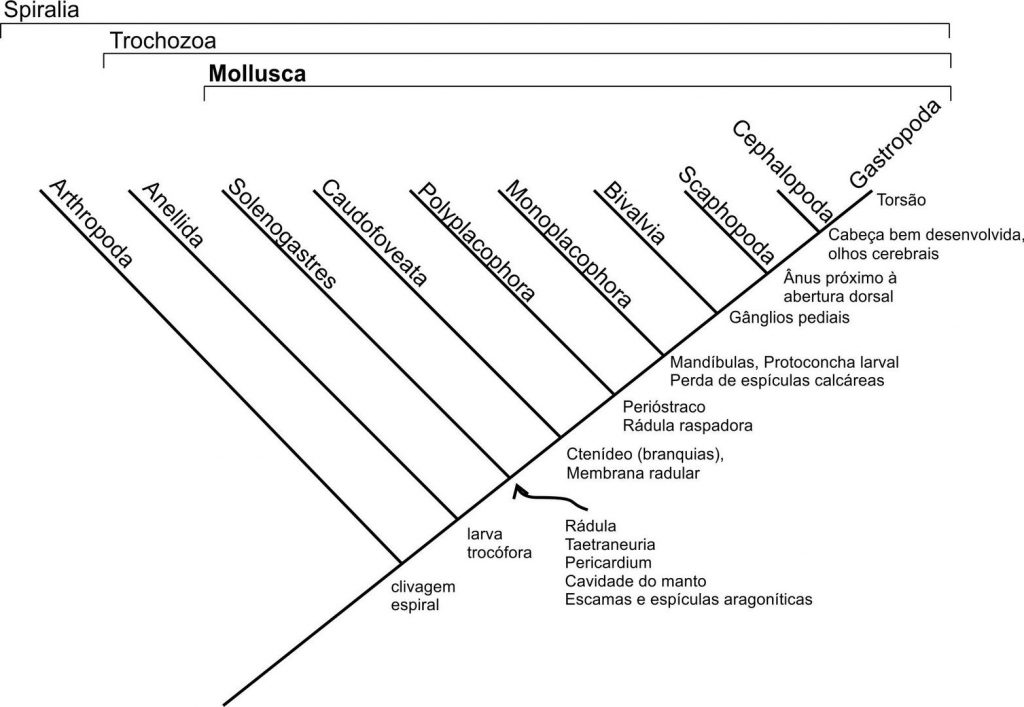
Figura 6.2. Uma dentre as muitas filogenias e caracteres que determina as relações internas dos diferentes clados de Mollusca (Brusca e Brusca, 1990).
A Revolução Cambriana do Substrato
Também chamada de Revolução Agronômica, a interpretação dos padrões de diversidade reconhecido nas sucessões de rochas cambrianas em todo o mundo, tem sido útil para a compreensão das causas que impulsionaram a acelerada disparidade (aparecimento de novos filos e classes) de metazoários em um curto intervalo de tempo durante o Período Cambriano. Em cerca de 5% do tempo de existência dos metazoários (algo em torno de 30 milhões de anos), 90% dos filos conhecidos apareceram no registro geológico, em rochas com idades entre 535 e 505 milhões de anos atrás. Conhecida como Explosão Cambriana, este que foi um dos eventos mais marcantes na história da vida, ainda busca por causas que expliquem o rápido aparecimento dos novos padrões morfológicos no registro cambriano.
A revolução cambriana do substrato está entre as diferentes possibilidades. Dentre seus principais sinais nas rochas está o aparecimento rápido e crescente de escavações verticais no substrato. Até então, as esteiras microbianas que revestiam o fundo oceânico impediam o acesso de invertebrados ao interior do substrato, ambiente no qual milhões de anos mais tarde evoluiria boa parte da diversidade de metazoários escavadores. A oportunidade apareceu com o surgimento de tecidos mineralizados. Aparatos raspadores de moluscos como Kimberella, a rádula, colaboraram com o rompimento das camadas microbianas que impermeabilizavam o substrato, oferecendo aos metazoários a oportunidade de acesso ao novo ambiente, e com ela novos modos de vida, alimentação, ancoragem e proteção.
Com o desaparecimento das esteiras microbianas das águas rasas oxigenadas, praticamente toda a vida macroscópica a ela associada, cerca de 200 gêneros dá diversificada Biota de Ediacara, foi extinta na primeira grande crise da vida multicelular.
O substrato anteriormente sulfuroso, anóxico e desabitado, tornou-se durante o Cambriano uma nova oportunidade para os experimentos evolutivos que logo ampliariam a diversidade de padrões corporais dentre os Metazoários. Embora outras causas que expliquem o desaparecimento das esteiras microbianas tenham sido propostas como, por exemplo, o atrito provocado pelas recém evoluídas carapaças rígidas, é certo que a radiação de moluscos dotados de rádula tenha tido forte impacto nas comunidades ediacaranas. Desde então, esteiras microbianas ocuparam ambientes hipersalinos ou alcalinos, impróprios à maioria dos metazoários.
A grande radiação de organismos escavadores dotados de estruturas mineralizadas, determinou o fim do grande Éon Proterozoico.
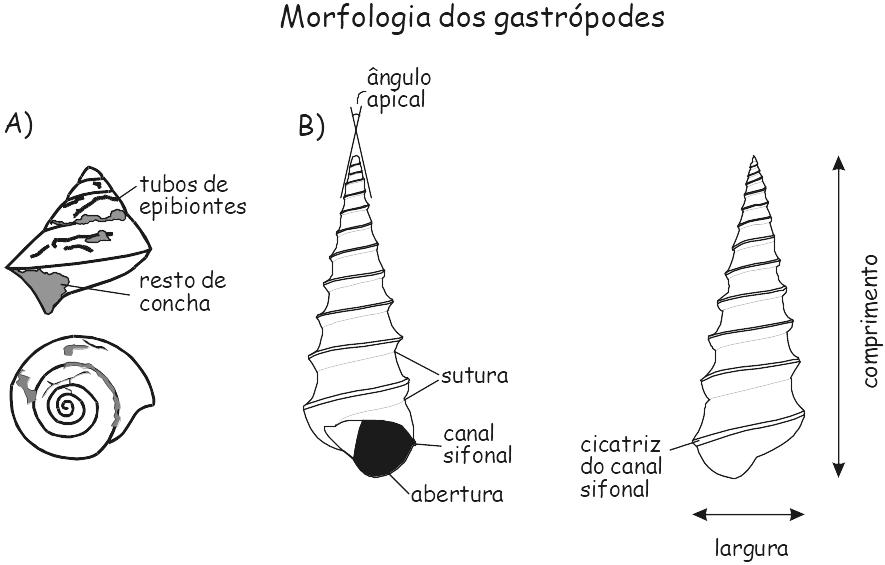
Figura 6.3. Morfologia dos gastrópodes. A) Euconospira sp. molde interno, Formação Piauí, (Carbonífero superior), Bacia do Paraíba, Brasil. B) Calyptera trochiformis, Eoceno.
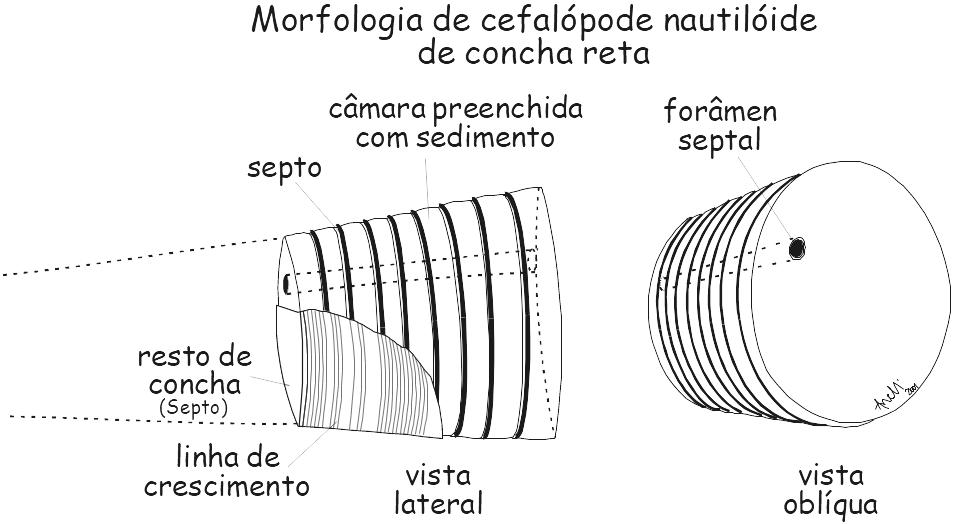
Figura 6.4. Morfologia dos nautilóides de concha reta. Orthoceras sp., (molde interno) Siluriano, Estados Unidos.
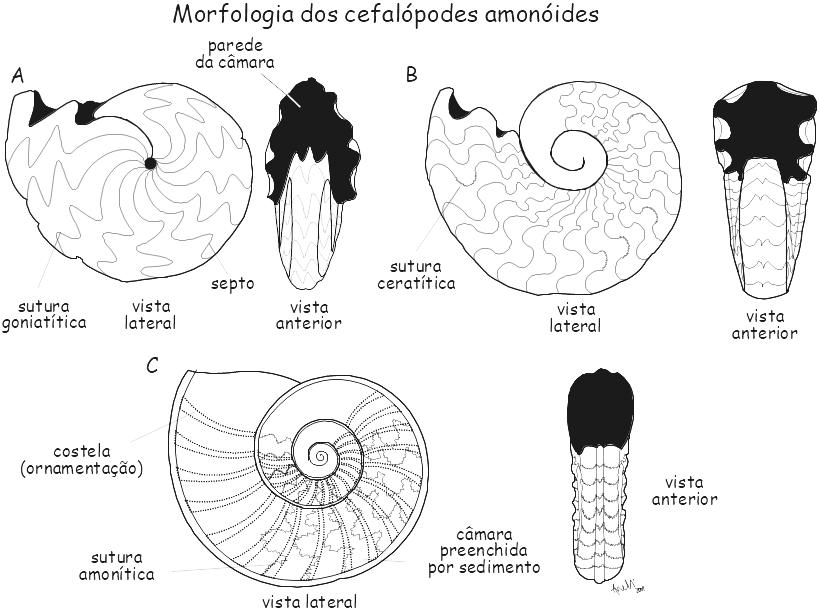
Figura 6.5. Morfologia dos amonóides de concha enrolada. A, Goniatites sp., Permiano, Austrália; B, Cearatites nodosus, Triássico médio, Alemanha; C, Asteroceras obtusun, Jurássico, Inglaterra.
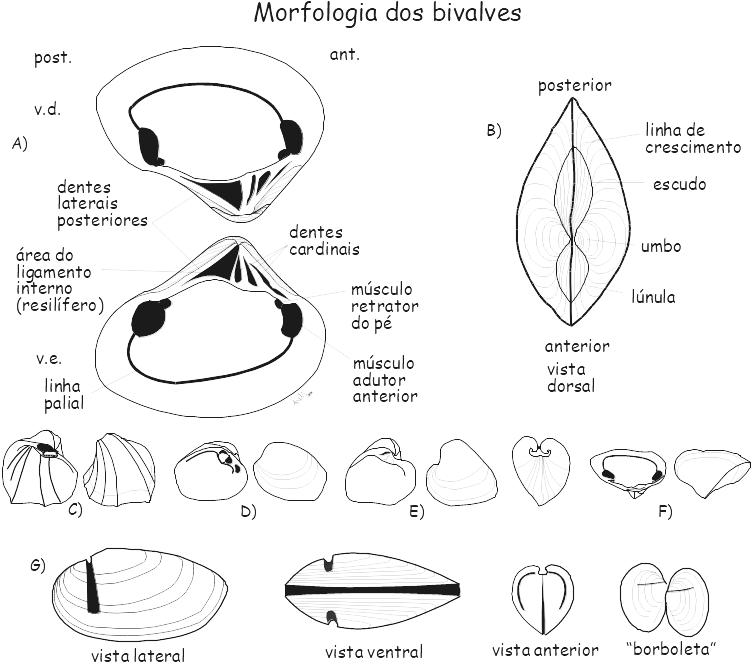
Figura 6.6. Morfologia interna e externa de concha de molusco bivalve.A-B Fucrassotella decipiens, Recente, Austrália. C-F Conchas silificadas. C Ferrasia cardinalis, valva esquerda; D Pinzonella ilusa, valva esquerda; E Plesiocyprinella carinata, (a) valva esquerda, (b) valvas conjugadas; F Jakezia brasiliensis, valva direita, Permiano, Formação Corumbataí, Bacia do Paraná; G Nuculites, molde interno, Formação Ponta Grossa (Devoniano), Bacia do Paraná.
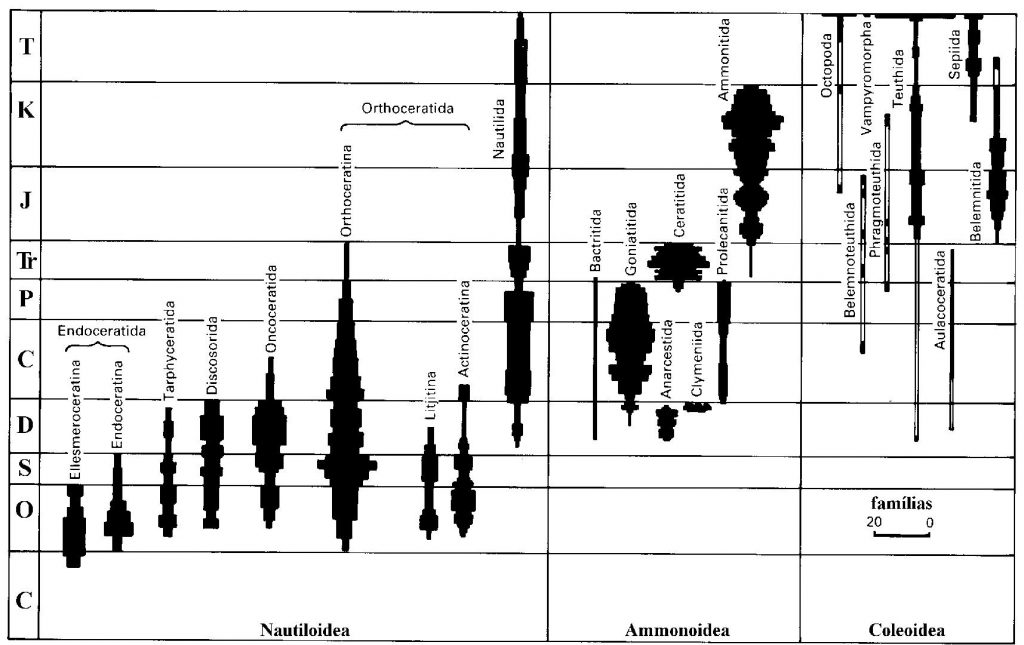
Figura 6.7. Distribuição geológica e abundâncias das três subclasses de cefalópodes (Clarkson, 1994).
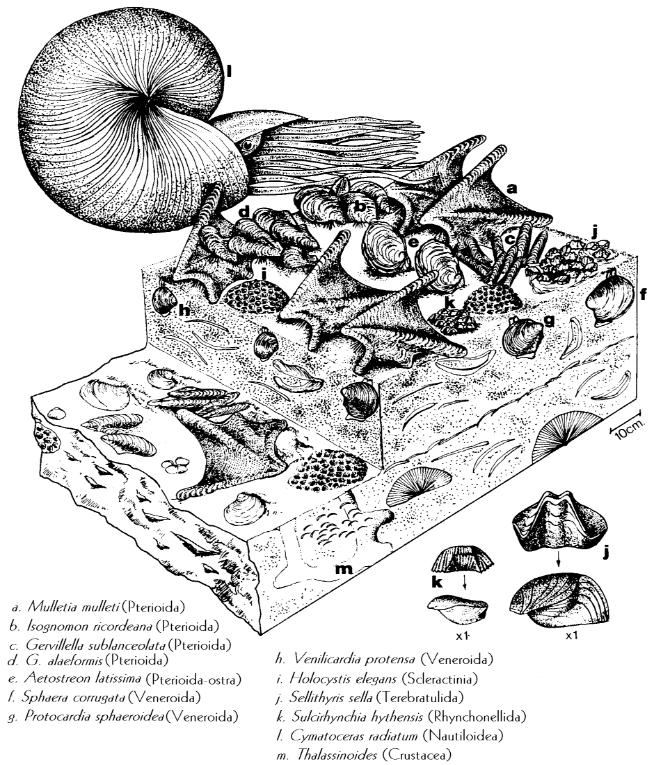
Figura 6.8. Reconstituição paleoecológica de uma comunidade jurássica de substrato marinho lamoso (McKerrow, 1978).
Copiado integralmente com a autorização dos autores:
Anelli, L.E.; Leme, J.M.; Oliveira, P.E.; Fairchild, T,R. 2020. Paleontologia. Guia de aulas práticas, uma introdução ao estudo dos fósseis. Universidade de São Paulo, Instituto de Geociências, 8a ed., 104p.
*Todos os direitos do textos e figuras são reservados aos autores