Galeria do Pré-Cambriano

















Introdução
Nos últimos anos, pesquisas em vários campos têm revelado diversos aspectos do surgimento e evolução precoce da vida na Terra. Bioquímicos têm simulado possíveis condições ambientais dos primórdios da Terra, na tentativa de entender como se formaram os compostos básicos para a origem da vida. Astrônomos e planetólogos investigam a atmosfera e superfície dos planetas e asteróides e procuram novos sistemas solares na busca de evidências de vida fora da Terra. Os geólogos procuram restos e vestígios de vida nas rochas sedimentares mais antigas da Terra, depositadas antes mesmo do aparecimento dos primeiros metazoários com carapaças e conchas, no Cambriano início do Éon Fanerozoico. São os paleontólogos, biogeoquímicos e estratígrafos que vasculham este registro pré-cambriano, à procura de fósseis que possam elucidar a origem e evolução inicial da vida.
O termo Pré-Cambriano refere-se ao imenso período de tempo que antecedeu o Cambriano, a primeira subdivisão da Era Paleozoico, que, juntamente com as eras Mesozoico e Cenozoico, faz parte do Éon Fanerozoico (de phaneros e zoos, “vida visível”). O Pré-Cambriano é subdividido em três éons, o Hadeano, o mais antigo, englobando o tempo desde a origem da Terra, há 4,56 Ga, (Giga-anos, ou bilhões de anos) até 3,85 Ga, que, excluindo uma ou duas ocorrências mais antigas, é a idade das rochas na base do registro geológico mundial; o Arqueano, de 3,85 Ga até 2,5 Ga; e o Proterozoico, o mais novo, de 2,5 Ga até 542 Ma (Mega-anos, ou milhões de anos) (Figura 2.1). Os geólogos ainda subdividem os éons Arqueano e Proterozoico em eras, designando-as por meio dos prefixos Paleo-, Meso-, e Neo- para as para os intervalos precoce, mediano e tardio, respectivamente, de cada éon.
Embora existam cristais do mineral zircão formados há 4,4 Ga presentes como grãos de areia em rochas arqueanas do SW da Austrália, as rochas mais antigas da Terra – os gnaisses de Acasta, no norte do Canadá – são bem mais novas, de “apenas” 4,03 Ga. Não existe, portanto, um registro geólogico do primeiro meio bilhão de anos da história da Terra. Durante este período tanto a Terra como a Lua e os outros planetas passaram por processos de estruturação interna, formação da litosfera (crosta e manto), no caso da Terra, surgimento da hidrosfera e atmosfera. Ao mesmo tempo, caíam sobre a superfície do planeta grande número de meteoritos, asteróides e cometas, que abundavam no Sistema Solar nesta época. Por isso, o Hadeano pode ser considerado a “Fase Cósmica ou Lunar” da nossa história.
Vida pré-cambriana
Se os primeiros metazoários com conchas e carapaças surgiram somente há cerca de 550 Ma, muitas inovações evolutivas devem ter se originado bem antes desta data. Logicamente, animais sem carapaças devem ter existido por um tempo indeterminado antes disto, e, inicialmente, devem ter sido minúsculos, provavelmente microscópicos. É evidente, também, que os animais (que são consumidores ou heterótrofos pluricelulares), devem ter sido precedidos por organismos produtores, ou seja, por autótrofos fotossintetizadores, inicialmente microscópicos (microalgas) evoluindo, posteriormente, para formas pluricelulares e macroscópicas (macroalgas).
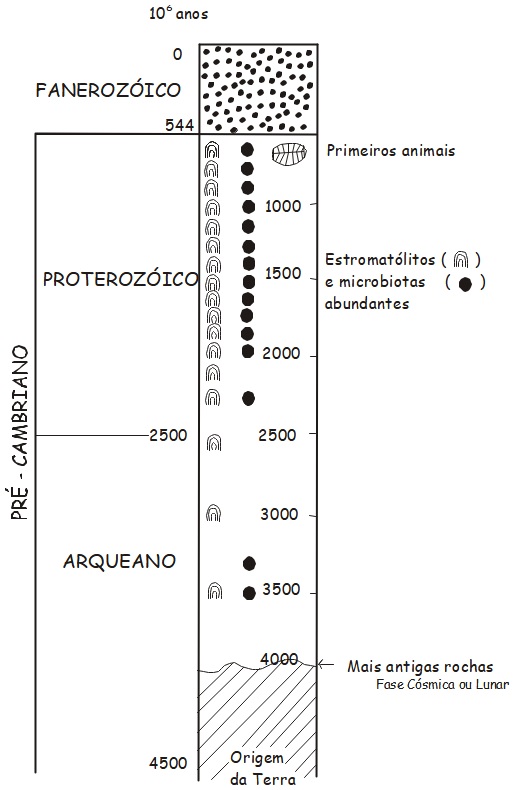
Figura 2.1. Tempo geológico e a distribuição de estromatólitos e microbiotas no Pré- Cambriano.
Mas algas e animais são idênticos em termos de organização intracelular: ambos são eucarióticos, isto é, providos de organelas intracelulares, núcleo e cromossomos lineares e capazes de se reproduzir assexuadamente (por mitose) e, na maioria dos casos, também sexuadamente (por meiose). Antes dos eucariontes surgirem, porém, deve ter existido um mundo só de procariontes, microrganismos sem núcleo e organelas intracelulares, e com cromossomos circulares e apenas formas simples de reprodução assexuada (fissão), como as bactérias, cianobactérias (“cianofíceas”) e arquebactérias atuais.
E antes dos procariontes o que teria existido? Evidentemente, apenas sistemas de vida ainda mais símples, como, por exemplo, meros complexos de proteínas e/ou ácidos nucléicos, desprovidos de qualquer invólucro celular, que se “alimentavam” de compostos orgânicos, produzidos abioticamente na hidrosfera e na atmosfera primitivas. Finalmente, o ponto de partida de toda a história biológica na Terra teria sido a própria origem da vida.
Fósseis pré-cambrianos
Embora estudos bioquímicos e experiências biológicas possam nos fornecer hipóteses sobre a origem e evolução precoce da vida, ainda são os fósseis que documentam e corroboram estas hipóteses e estabelecem a ordem e cronologia dos eventos evolutivos. Para Darwin, a aparente ausência de fósseis em rochas pré-cambrianas constituiu um problema sério da Paleontologia. Contrastado com o registro extremamente abundante de fósseis fanerozoicos, o registro pré-cambriano parecia paupérrimo, aparentemente sem evidências dos organismos precursores dos primeiros invertebrados conchíferos, que aparecem em grande abundância e diversidade logo no início (período Cambriano) do Fanerozoico.
Sabemos agora que esta visão do registro fóssil estava errada. Primeiro, pelas evidências acumuladas desde 1960, reconhecemos uma verdadeira explosão adaptativa dos metazoários macroscópicos nas poucas dezenas de milhões de anos que antecederam o Fanerozoico. Os invertebrados macroscópicos não gozavam, necessariamente, de uma longa história pré-cambriana. E, segundo, descobriu-se um registro fóssil pré-cambriano surpreendentemente rico e extenso, não de organismos macroscópicos, mas sim de formas microscópicas, em grande parte atribuíveis aos procariontes unicelulares e coloniais. Dessa forma, o Pré-Cambriano pode ser apelidado de era da vida microscópica ou da vida procariótica, justificando o termo Criptozoico (“vida escondida”), usado antigamente para o Pré-Cambriano mas agora considerado arcaico.
Fósseis pré-cambrianos são, via de regra, de seis tipos distintos, separados em duas categorias:
Vestígios:
- Impressões (moldes) de invertebrados primitivos, sem carapaças, restritos ao fim do Proterozoico
- Icnofósseis, raros e restritos ao fim do Proterozoico
- Estromatólitos, muito comuns em rochas carbonáticas (calcários) pré-cambrianas
Restos:
4) Microfósseis orgânicos, permineralizados por sílica em calcários, ou compactados e
incarbonizados em folhelhos
5) Fósseis químicos, evidências orgânicas (compostos ou relações isotópicas de Carbono)
de origem biológica, resistentes aos processos geológicos (p.ex, carvão, petróleo etc.)
6) Partes esqueléticas inorgânicas, (conchas, carapaças, espículas etc., de calcita, fosfato ou sílica), restritas ao fim do Proterozoico
Estromatólitos
Estromatólitos são estruturas biossedimentares, de laminação geralmente fina e convexa, que resultam do crescimento e das atividades metabólicas de comunidades de micróbios bentônicos autotróficos, sobremaneira de cianobactérias (antigamente conhecidas como cianofíceas ou “algas azúis”). O crescimento rápido de microrganismos filamentosos e, subordinadamente, de colônias de células cocóides, acompanhado pela secreção de abundante mucilagem, fixa estas comunidades firmamente no substrato e muitas vezes aglutina ou retém partículas finas de sedimento, na sua superfície, formando verdadeiras “esteiras microbianas” de poucos milímetros de espessura. O empilhamento de sucessivas esteiras e sua consolidação (litificação) pela precipitação precoce de carbonato de cálcio resultam em estruturas maiores — estromatólitos – de diversas formas (Fig. 2.2). A grande maioria dos estromatólitos se formou em ambientes litorâneos, desde a supramaré até a inframaré, mas há também muitas ocorrências de estromatólitos em ambientes continentais (lagos, rios, fontes térmicas etc.).
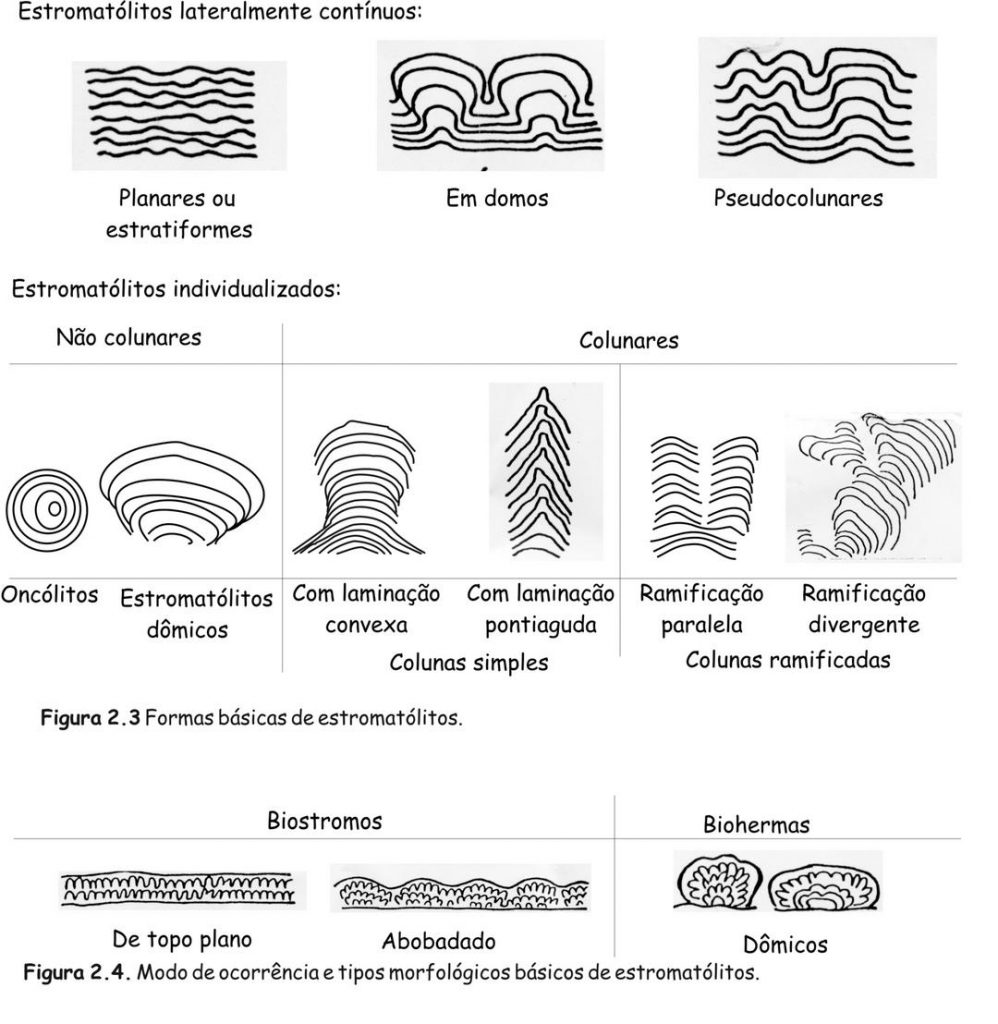
Os estromatólitos podem apresentar morfologia estratiforme, dômica, colunar, pseudocolunar ou até esferóide (estes são chamados de oncóides ou oncólitos). Na década de 60, quando se iniciaram estudos de estromatólitos atuais, pensava-se que os eles pudessem ser caracterizados, morfologicamente, por fórmulas simples, baseadas no formato e empilhamento de suas lâminas em combinação com o grau de ligação entre indivíduos próximos, como ilustrado na Fig. 2.2. Embora este esquema permita descrever estromatólitos simétricos e símples, é pouco prático para formas mais complexas, que, aliás, são muito comuns.
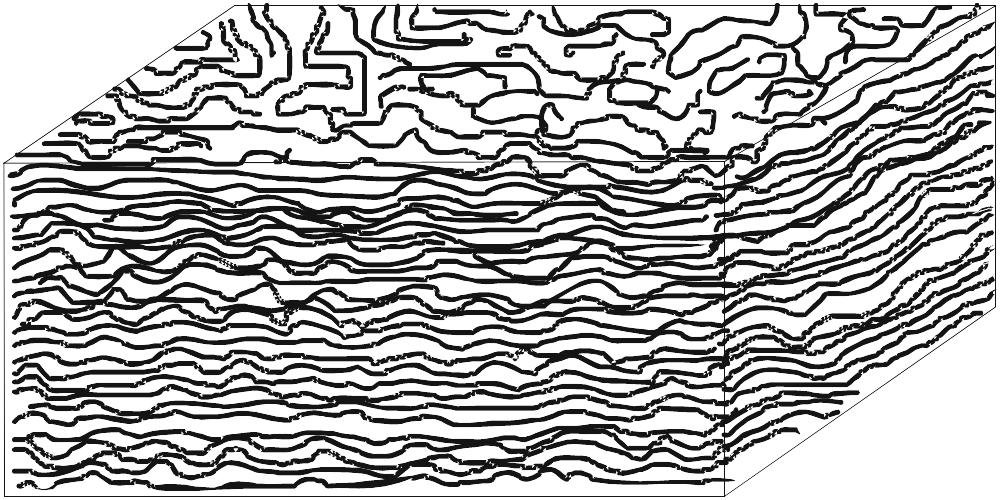
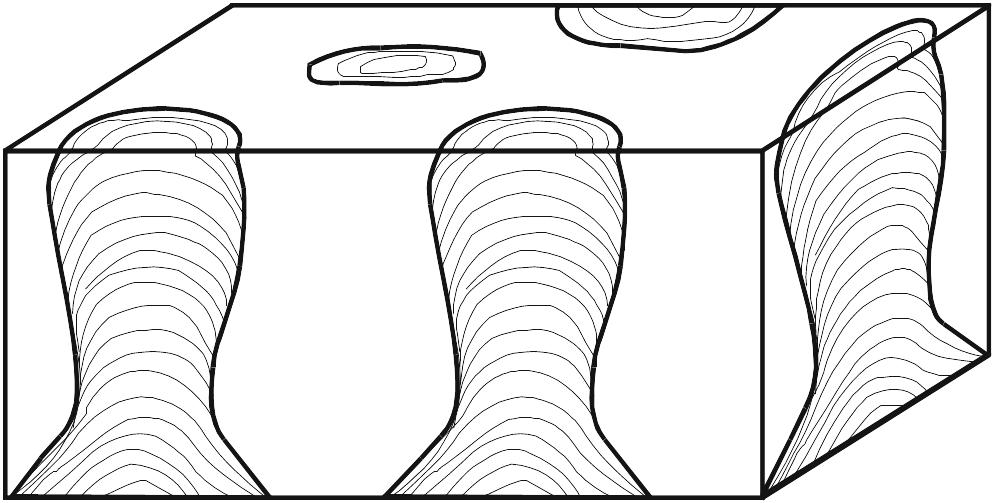
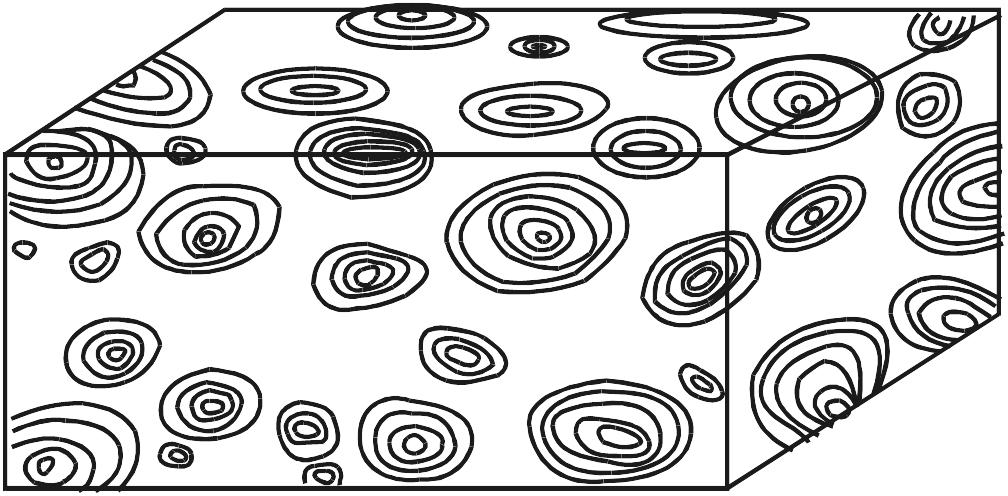
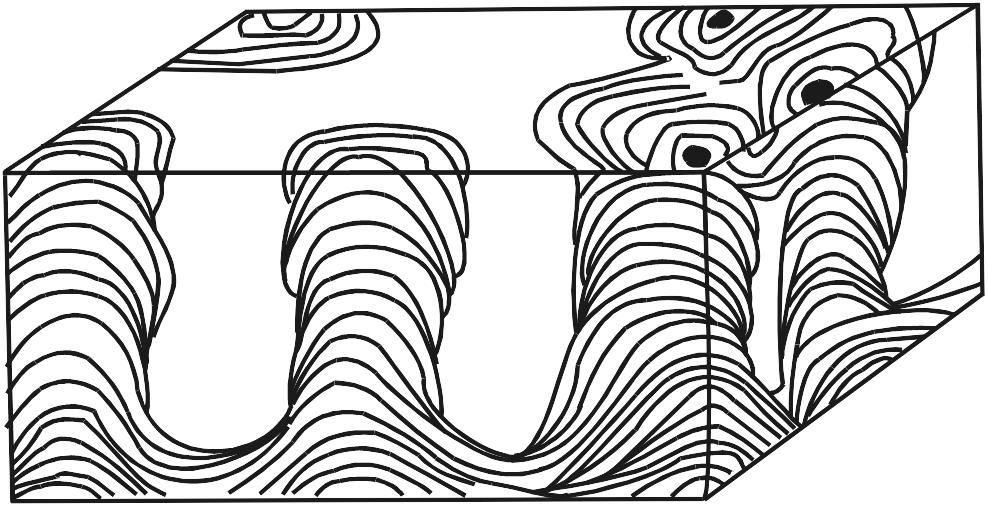
Figura 2.2. a-d. Formas básicas de estromatólitos simples, segundo Logan, Rezak e Ginsburg (1964). LLH = Laterally Linked Hemispheroids; SH = Stacked Hemispheroids; SS = Stacked Spheroids. Em d, as formas LLH podem dar origem a formas SH (e vice-versa).
Atualmente, portanto, as feições morfológicas mais úteis para a descrição de estromatólitos são: 1) forma geral (Fig. 2.3); 2) dimensões (altura, largura e comprimento, ou diâmetro); 3) características da laminação (grau de convexidade, espessura, relevo laminar, regularidade).
Estromatólitos variam em tamanho de milimétricos a métricos e ocorrem isolados ou densamente agrupados em corpos lenticulares chamados biohermas, ou em camadas mais ou menos regulares conhecidas como biostromas (Fig. 2.4). Os mais antigos (Austrália e África do Sul) datam de 3,5 Ga, mas também existem formas atuais. Escassos no Arqueano e extremamente abundantes no Proterozoico, os estromatólitos tornam-se restritos no Fanerozoico como conseqüência da expansão dos animais e plantas. Estromatólitos são conhecidos em rochas pré-cambrianas de todos os continentes, menos Antártica.
No Brasil, os estromatólitos mais antigos ocorrem no Quadrilátero Ferrífero, perto de Ouro Preto, MG, e têm idade entre 2,1 e 2,4 Ga, mas apresentam-se deformados e recristalizados. O principal registro de estromatólitos no País é nos terrenos meso- e neoproterozoicos dos estados de GO, MG, PR, BA (onde ocorrem formas substituídas por fosfato) e no DF; também foram registrados em SP, MS, MT, PA, AM e CE.
Estromatólitos também ocorrem no Fanerozoico do Brasil, nas formações permianas Corumbataí/Estrada Nova (SP, PR) e Pedra de Fogo (MA), bem como em sedimentos quaternários da Lagoa Salgada (agora praticamente seca), no litoral norte do Estado do Rio de Janeiro, onde se formaram há menos de 2.000 anos. Tipicamente, estromatólitos fanerozoicos formaram-se em ambientes restritos, comumente hipersalinos, evidentemente inóspitos tanto à maioria de organismos fotossintetizantes, concorrentes das cianobactérias, como aos organismos heterotróficos (como gastrópodes), que se alimentam das comunidades microbianas.
A importância dos estromatólitos na Paleontologia e Geologia.
Figs. 2.5 e 2.6 ilustram, esquematicamente, a distribuição de estromatólitos modernos em Hamelin Pool, no extremo sul da Shark Bay, NW da Austrália. Observe na Fig. 2.5 a correlação entre a energia do ambiente e a forma dos estromatólitos. Assim, os estromatólitos estratiformes (LLH) se formam na zona de menor energia, enquanto as formas colunares (SH) e oncólitos (SS) se formam em zonas agitadas.
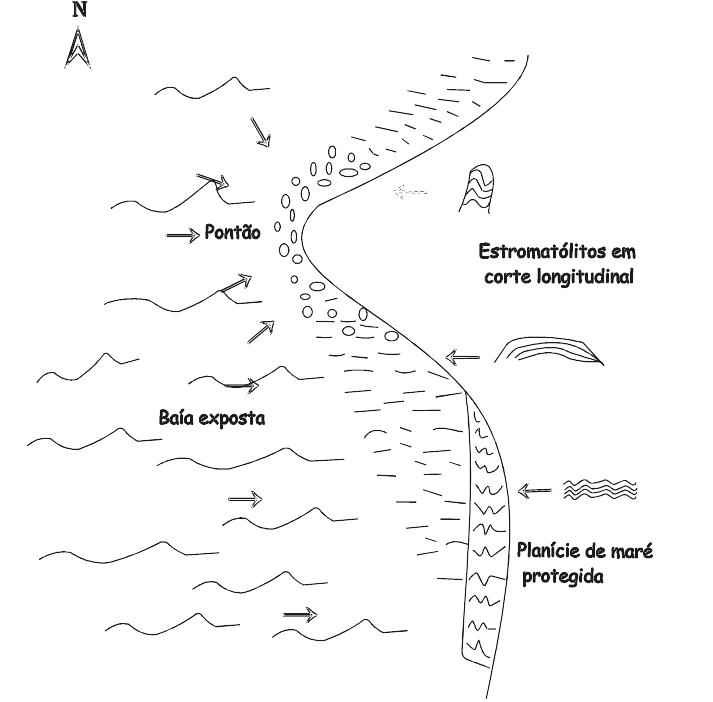
Fig. 2.6 mostra outro aspecto da relação energia/forma, ou seja, a tendência dos estromatólitos ficarem mais cilíndricos onde as ondas e correntes vêm de várias direções (nos pontões, por exemplo), e alongados, horizontalmente, onde a direção das correntes é constante (na baia exposta).
Por que a grande maioria de estromatólitos são construídos por organismos fotossintetizadores, podemos fazer as seguintes inferências importantes:
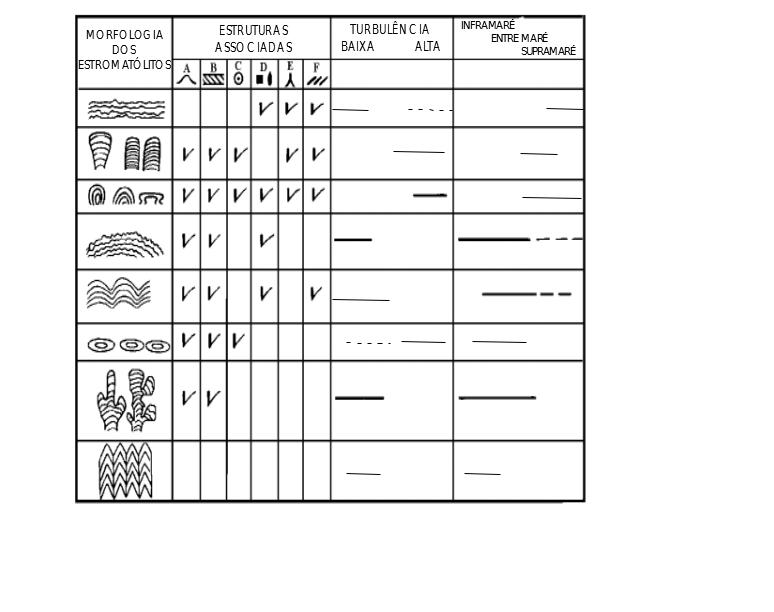
a) A rocha portadora deve ter se formado dentro da zona fótica, ou seja, a profundidades rasas, certamente a menos de 200 m, e, no caso de biohermas e biostromas bem desenvolvidos, provavelmente, desde poços na zona de supramaré até profundidades de poucas dezenas de metros. A análise da forma de estromatólitos e das estruturas sedimentares associadas permite inferir as condições hidrodinâmicas e a profundidade relativa do ambiente deposicional (Tabela 2.1).
b) Sendo os estromatólitos indicadores de águas rasas, eles podem ajudar na reconstituição da paleogeografia da linha da costa da época.
c) As lâminas estromatolíticas são sempre convexas para cima, o que permite ao geólogo de campo determinar topo e a base em seqüências rochas deformadas.
Os estromatólitos são, afinal, uma importante evidência de vida, não apenas de microrganismos fotossintetizadores, mas, também, de ecossistemas bentônicos razoavelmente complexos. Daí, a presença de estromatólitos em rochas de 3,5 Ga de idade na Austrália e África do Sul constitui uma das mais antigas evidências de vida conhecidas.
Finalmente, pesquisadores na antiga URSS demonstraram, empiricamente, que se pode caracterizar intervalos do éon Proterozoico, da ordem de várias centenas de milhões de anos, com base nas sucessão de associações de estromatólitos distintas e, assim, utilizar estromatólitos, em certos casos, para correlação temporal de sucessões de rochas distantes entre si. Ao contrário da bem conhecida sucessão evolutiva de faunas e floras no Fanerozoico, que constitui a base da correlação bioestratigráfica e da escala de tempo geológico, as diferenças constatadas entre associações estromatolíticas não obedecem hierarquia evolutiva de caracteres morfológicos geneticamente controlados. Os estromatólitos, afinal, são estruturas biossedimentares, o produto da interação de comunidades microbianas bentônicas com o meio ambiente. Portanto, não se pode falar de formas estromatolíticas “ancestrais” e “descendentes”, ou formas “primitivas” e “avançadas”. Já foi sugerido que as diferenças observadas talvez reflitam mudanças paleogeográficas na forma e distribuição dos continentes e oceanos ao longo do tempo, que teriam resultado na reorganização, em escala global, de comunidades microbianas bentônicas.
Registro paleontológico e geológico dos principais eventos evolutivos do Pré-Cambriano
Nem todos os eventos evolutivos importantes ocorridos no Pré-Cambriano deixaram um registro claro nas rochas. Por exemplo, não há nenhuma evidência geológica palpável da origem da vida. E mesmo se existisse, provavelmente passaria despercebida por nos, dada sua simplicidade, fragilidade e tamanho ínfimo. Contudo, a presença, em algumas das mais antigas rochas sedimentares conhecidas, de estromatólitos, supostos microfósseis cianobacterianos (Fig. 2.7) e possíveis fósseis químicos indicativos de fotoautotrofismo implica no surgimento da vida e dos primeiros procariontes, processos de fotossíntese e ecossistemas bentônicos complexos antes de 3,5 Ga atrás. Agora, uma vez que o intenso bombardeio meteorítico sofrido pela Terra entre 4,5 e 3,8 Ga certamente teria dificultado a sobrevivência de qualquer forma de vida terrestre no éon Hadeano, abre-se a possibilidade de que a origem da vida, como nós a conhecemos, e de organismos procarióticos, fotossíntese e ecossistemas microbianos variados, tenha ocorrido no pequeno intervalo de tempo entre 3,8 e 3,5 Ga!
Toda teoria sobre a origem da vida condiciona seu aparecimento a um ambiente primevo de caráter químico redutor, para assegurar que os compostos orgânicos simples, necessários para a vida, não se oxidassem. Assim, a ascensão de oxigênio como gás estável em nossa atmosfera só pode ter ocorrido muito após a origem da vida. De fato, o oxigênio atmosférico é de origem biológico, produto da fotossíntese, que transforma água e dióxido de carbono em matéria orgânica e libera oxigênio. Esta reação é reversível, de modo que o oxigênio é consumido na decomposição da matéria orgânica. . . . a não ser que a matéria orgânica seja soterrado e assim isolada, ou “seqüestrada”, nos sedimentos, longe do alcance do oxigênio. O oxigênio que “sobra” na superfície passa, então, a oxidar outras coisas, por exemplo, gases vulcânicos, rochas expostas e compostos dissolvidos na hidrosfera, como ferro ferroso. Com a expansão das cianobactérias nos oceanos e o acúmulo de cada vez mais matéria orgânica nas rochas, procedeu-se à oxidação mais ou menos completa das terras expostas e oceanos, permitindo, em seguida, o acúmulo progressivo de oxigênio na atmosfera.
O oxigênio foi importante, biologicamente, porque condicionou o aparecimento do processo de respiração, um processo metabólico que produz ATP mais eficientemente do que a fermentação, quando o nível de O2 supera 1% do nível atualmente presente na atmosfera. Os eucariontes se aproveitaram desse processo e acabaram imprimindo um novo ritmo à evolução.
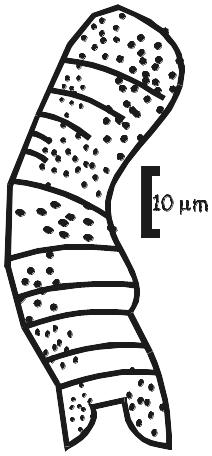

Figura 2.7. Desenhos interpretativos de microfósseis carbonosos (A-B) observados em lâminas petrográficas de sílex da Formação Apex (3,5 Ga), NW Austrália. Escala de 10µm em B serve para todas as figuras. (Adaptado de Schopf, 1993).
O oxigênio também condicionou a formação da camada de ozônio (O3) na atmosfera, que absorve a faixa de luz ultravioleta mais danosa a vida. Sem oxigênio na atmosfera,raios ultravioletas nocivos penetrariam até 10 m de profundidade nos oceanos; com 1% do nível atual de oxigênio, penetrariam 30 cm; e sómente quando o oxigênio atingisse 10% do seu nível atmosférico atual, formar-se-ia uma camada de ozônio capaz de proteger a vida dentro e até fora d’água.
Evidências importantes na história do oxigênio na atmosfera são fornecidas pela Geologia e Paleontologia. Primeiro, a grande maioria dos depósitos de ferro no mundo, como os do Quadrilátero Ferrífero e da Serra dos Carajás depositou-se, por processos sedimentares, no período de 3,0 a 2,0 Ga, e só pode ser explicado se pressupormos condições anóxicas para as regiões profundas dos oceanos dessa época. Microfósseis em rochas lateralmente equivalentes (Fig. 2.8) parecem representar apenas de formas procarióticas, inclusive algumas bem bizarras. Certamente, antes de 2,0 Ga, o nível de oxigênio deve ter sido muito baixo.

Próximo do fim da deposição dos grandes depósitos de ferro, apareceram em grande escala, pela primeira vez, rochas sedimentares continentais depositadas sob condições claramente oxidantes, conhecidas como red beds, ou “camadas avermelhadas”, em alusão à coloração imposta à rocha pelo óxido de ferro. Nessa mesma época, em torno de 2 Ga atrás, aparecem os primeiros fósseis de supostos eucariontes: a) microfósseis unicelulares muito maiores do que procariontes modernos b) biomarcadores químicos (fósseis químicos) indicativos de compostos orgânicos produzidos somente por eucariontes, e c) impressões em forma de uma “mola achatada”, visíveis ao olho nu, denominadas de Grypania spiralis, e interpretadas por alguns como possível macroalga primitiva. Uma vez que praticamente todos os eucariontes utilizam-se da respiração e sabendo-se que este processo só é eficiente em ambientes com mais de 1% do nível atual de oxigênio, estes fósseis sugerem que este patamar deve ter sido ultrapassado em torno de 2,0 Ga atrás.
Figura 2.8. Microfósseis carbonosos em estromatólitos permineralizados por sílica da Formação Gunflint (2,1 Ga), SE Ontário, Canadá. Parte A aumentada cerca de 1000X; partes B-D, cerca de 2000x. Observe a esteira microbiana formada por filamentos com cocóides subordinados em B e C. retratam formas bizarras. O estudo desta microflora, publicado em 1965 por Barghoorn & Tyler, marcou o início das investigações paleontológicas modernas de rochas pré-cambrianas.

De 2,0 Ga até cerca de 1,0 Ga, microfósseis eucarióticos tornaram-se cada vez mais comuns, mas, mesmo assim, continuaram morfologicamente muito simples, distinguíveis dos procarióticos principalmente por seu tamanho maior. Neste mesmo intervalo apareceram incarbonizações e impressões de tamanho milimétrico a centimétrico sob forma de fitas e elipses, interpretadas como posssíveis talos as macroalgas.
É só no Neoproterozoico (1,0-0,54 Ga), contudo, que aparecem evidências seguras e abundantes da diversificação dos eucariontes, talvez como resultado do surgimento da reprodução sexuada, cuja evidência mais antiga, algas vermelhas microscópicas do Canadá, data de 1,2 Ga. Estas evidências incluem microfósseis ornamentados (Fig. 2.9), possíveis algas calcárias e, ao final do Neoproterozoico (entre 600 e 542 Ma), as primeiras evidências de metazoários macroscópicos sob a forma de a) icnofósseis, inicialmente muito símples, tornando-se mais complexos e variados pouco antes da passagem para o Cambriano, b) impressões de invertebrados desprovidos de carapaças, como as de Ediacara, no sul da Austrália (Fig. 2.10) que constituem a mais antiga fauna fóssil conhecida até, e c) na “véspera do Cambriano”, os primeiros fósseis com conchas de carbonato de cálcio, uma evidência de organismos capazes de secretar compostos inorgânicos (biomineralização), como Cloudina lucianoi, da região de Corumbá, MS (Fig. 2.11).
Voltando à história do oxigênico atmosférico, estudiosos, levando em conta a forma muito achatada dos fósseis da fauna de Ediacara, calcularam que os organismos originais não poderiam ter sobrevivido se a atmosfera não contivesse de 6 a 8% do seu nível atual de oxigênio.
Finalmente, no Fanerozoico, com o desenvolvimento das primeiras florestas no fim do Paleozoico, há 350-250 Ma, o nível de oxigênio atmosférico deveria ter dado novo salto, chegando, seguramente, a níveis iguais ao atual (ou até maiores) no Mesozoico, entre 200 e 100 Ma atrás, quando da origem e rápida expansão dos grandes répteis (dinossauros) e das angiospermas.
Copiado integralmente com a autorização dos autores:
Anelli, L.E.; Leme, J.M.; Oliveira, P.E.; Fairchild, T,R. 2020. Paleontologia. Guia de aulas práticas, uma introdução ao estudo dos fósseis. Universidade de São Paulo, Instituto de Geociências, 8a ed., 104p.
*Todos os direitos do textos e figuras são reservados aos autores